Temporal Summation: How Small Signals Create Big Ideas
- Introduction and Definition of Temporal Summation
- The Neural Basis: Postsynaptic Potentials (PSPs)
- Distinction from Spatial Summation
- The Mechanisms of Persistence and Time Constants
- Role in Neural Integration and Decision Making
- Experimental Evidence and Measurement Techniques
- Clinical Relevance and Pathophysiology
Introduction and Definition of Temporal Summation
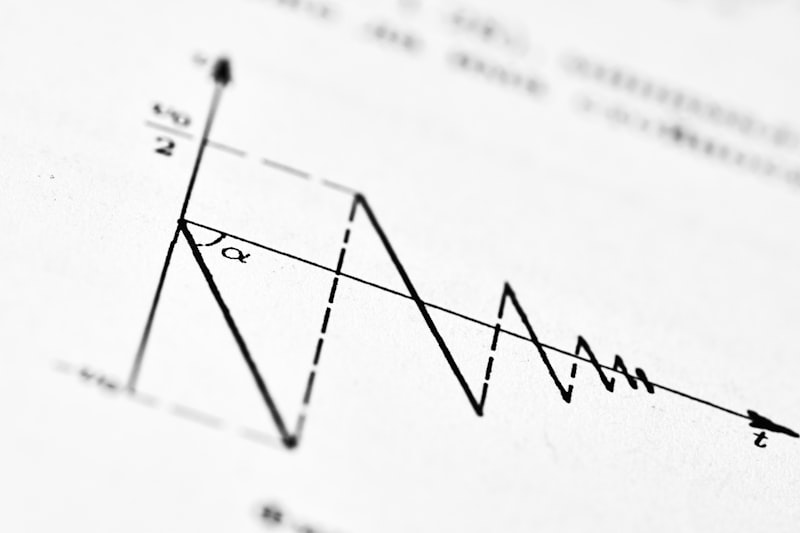
Temporal summation represents a critical neural mechanism fundamental to the integrative capacity of the nervous system, describing the process by which a neuron combines the effects of multiple, subthreshold synaptic inputs arriving sequentially at the same synapse over a short period of time. This sophisticated integration is essential for converting individually weak electrical signals into a powerful, cumulative depolarization capable of reaching the firing threshold and generating an action potential. The concept is predicated on the biophysical reality that a change in the postsynaptic membrane potential does not dissipate instantaneously. When a presynaptic neuron releases a neurotransmitter, causing a partial depolarization—known as a subthreshold excitatory postsynaptic potential (EPSP)—this voltage change persists for several milliseconds, governed by the membrane’s time constant. If a second EPSP arrives before the membrane potential has fully returned to its resting state, the residual electrical effect of the first potential adds algebraically to the second. This additive impact ensures that while neither potential alone is sufficient to elicit a reaction, their rapid succession results in an above-threshold depolarization, effectively inducing neural communication downstream.
The efficacy of temporal summation is directly proportional to the frequency of the incoming signals. A high-frequency burst of action potentials from the presynaptic terminal maximizes the chances that each subsequent neurotransmitter release will occur precisely during the lingering depolarization caused by the previous event. This sequential stacking of potentials allows the postsynaptic neuron to filter out random, low-frequency electrical noise while amplifying coherent, repeated signals that carry significant physiological information. Temporal summation thereby ensures that the nervous system is highly responsive to sustained or intense stimuli, even when the individual synaptic contribution is modest. This mechanism underscores the neuron’s role not as a simple switch, but as a complex integrator, calculating the weighted average of incoming electrical activity across time to make a decision about whether or not to fire.
The mathematical basis of this process is governed by the principles of cable theory, which describe how electrical signals propagate and decay within dendrites. The duration of the postsynaptic potential is significantly longer than the transient opening of the initial ion channels, leading to a window of opportunity for summation. Without this capacity for temporal integration, the sheer number of weak synapses in the brain would render much of the complex signaling ineffective. Temporal summation is thus indispensable for transforming the discrete, rapid-fire events of presynaptic spiking into the sustained, graded potentials necessary for reliable and robust signal propagation across neural circuits, forming the basis of coordinated perception and response.
The Neural Basis: Postsynaptic Potentials (PSPs)
To understand the mechanics of temporal summation, a deep appreciation of the nature of the postsynaptic potential (PSP) is required. PSPs are localized, graded potentials, meaning their amplitude is not fixed but varies continuously based on the amount of neurotransmitter released and the number of receptor channels activated. Unlike the all-or-nothing nature of the action potential, PSPs are analog signals that contribute incrementally to the overall membrane voltage. They are categorized primarily as either Excitatory Postsynaptic Potentials (EPSPs), which depolarize the cell (moving the potential closer to the threshold), or Inhibitory Postsynaptic Potentials (IPSPs), which hyperpolarize the cell (moving the potential further away from the threshold). Temporal summation operates on both types. A rapid succession of EPSPs results in cumulative depolarization, whereas a rapid succession of IPSPs results in cumulative hyperpolarization, stabilizing the membrane potential and actively suppressing excitability.
The critical feature enabling temporal summation is the relatively slow decay phase of the PSP. When excitatory neurotransmitters, such as glutamate, bind to their receptors, ligand-gated ion channels open, typically allowing the influx of positive ions like sodium. This inflow constitutes the rising phase of the EPSP. However, even after the neurotransmitter is rapidly cleared from the synaptic cleft via reuptake or enzymatic degradation, the resulting change in membrane voltage persists. The membrane potential does not instantly snap back to its resting state; rather, it decays exponentially over a period of milliseconds. This decay is governed by the inherent resistance and capacitance of the neuronal membrane. Because the typical action potential threshold requires a significant depolarization (often 10–20 mV) and a single EPSP rarely contributes more than 1 mV, the necessity of accumulating multiple subthreshold events becomes biologically mandatory.
The location of the synapse on the dendritic tree also subtly influences the efficiency of temporal summation. Although the summing process occurs locally at the synapse, the accumulated potential must then passively spread through the dendritic branches to the axon hillock, the neuron’s integration center. During this passive spread, the signal inevitably suffers attenuation due to internal resistance and membrane leakage. Therefore, synapses located on proximal dendrites or directly on the soma have a greater influence on the final integration outcome than those located distally. However, for temporal summation to be effective, the interval between successive PSPs must be short enough to overcome both the natural decay rate (time constant) and the spatial decay (length constant) incurred during propagation toward the axon hillock. This interplay of temporal and spatial factors dictates the precise computational capability of any given neuron.
Distinction from Spatial Summation
While both temporal summation and spatial summation are mechanisms of neural integration that determine whether a neuron fires an action potential, they are differentiated by the physical arrangement and timing of the incoming synaptic inputs. Temporal summation focuses on the intensity of a single input source over time, requiring rapid, sequential firing from one presynaptic terminal. In contrast, spatial summation involves the simultaneous integration of postsynaptic potentials originating from multiple distinct synapses located at various points across the dendritic tree and the soma of the postsynaptic neuron. These geographically separate inputs converge on the integration zone, and their algebraic sum—taking into account both excitatory (depolarizing) and inhibitory (hyperpolarizing) contributions—determines the outcome.
The key distinguishing factor is the input source and alignment. In temporal summation, the inputs are sequential and arrive from the same, highly active presynaptic source, demanding high-frequency firing. In spatial summation, the inputs are concurrent, arriving simultaneously from multiple, independent presynaptic sources. For example, a neuron might receive an EPSP from a synapse on a distal dendrite and simultaneously an IPSP from a synapse near the soma. Spatial summation combines these two opposing forces instantaneously. If the depolarization outweighs the hyperpolarization, the net result moves the neuron closer to threshold. If the hyperpolarization dominates, the neuron is inhibited.
In the physiological reality of the central nervous system, neurons rarely rely exclusively on one form of summation. The typical postsynaptic neuron is subjected to a constant barrage of thousands of signals arriving from diverse sources at varying frequencies. The true decision to fire an action potential involves a complex, ongoing computational process where the neuron continuously weighs the combined effects of both temporal summation (high-frequency inputs being amplified) and spatial summation (multiple sources converging). This combined integration ensures that the neuron fires only when there is sufficient evidence—both in terms of intensity (temporal summation) and breadth (spatial summation)—to warrant the propagation of the signal, thereby acting as a powerful filter and decision-making unit within the neural circuit.
The Mechanisms of Persistence and Time Constants
The physical property underpinning the efficacy of temporal summation is the membrane time constant ($tau$), which quantifies the speed at which the membrane potential responds to a change in current, and, crucially, the speed at which it returns to its resting state. A longer time constant signifies that the electrical effects of a PSP persist for a greater duration, providing an extended window during which subsequent PSPs can accumulate their effects. The time constant is mathematically defined as the product of the membrane resistance ($R_m$) and the membrane capacitance ($C_m$). High membrane resistance, which resists the leakage of ions back across the membrane, and high membrane capacitance, which relates to the membrane’s ability to store charge, both contribute to a longer time constant and enhanced temporal summation capability.
For effective temporal summation, the inter-stimulus interval (ISI) between successive presynaptic action potentials must be significantly shorter than the postsynaptic membrane time constant. If the ISI is short, the second PSP arrives while the membrane is still significantly depolarized from the first event, leading to a robust additive effect. If the ISI is long, the initial depolarization will have decayed back toward the resting potential, minimizing or eliminating the summative effect. Neurons are highly specialized; those involved in rapid processing, such as certain relay neurons, may possess short time constants to prioritize speed and prevent temporal blurring of signals. Conversely, neurons involved in complex integrative functions, such as those in the hippocampus or cortex, often exhibit longer time constants, enhancing their ability to integrate information over slightly longer durations necessary for pattern recognition and memory formation.
Furthermore, the time constant is not a static property but is dynamically regulated by the physiological state of the neuron. Neuromodulators and internal signaling pathways can influence membrane resistance by altering the activity of voltage-gated or leak ion channels. For example, the closure of potassium leak channels increases the membrane resistance, lengthens the time constant, and enhances the neuron’s overall excitability and capacity for temporal summation. This dynamic modulation allows the neuron to adjust its integration properties based on context, potentially increasing sensitivity during periods of arousal or attention. The time constant is thus a critical determinant of neuronal computational capacity, directly linking biophysical properties to functional signal processing.
Role in Neural Integration and Decision Making
Temporal summation plays a pivotal role in neural integration, serving as a primary mechanism by which the intensity and coherence of incoming signals are translated into functional neural output. In biological systems, the frequency of action potentials often encodes the magnitude or significance of a stimulus. A sensory receptor responding weakly might fire once, producing an isolated, subthreshold EPSP that is filtered out. However, a strong stimulus—such as a commanding visual cue or intense auditory input—will trigger a high-frequency barrage of presynaptic firing. Temporal summation ensures that only this high-frequency, patterned input is amplified sufficiently to cross the threshold, thereby providing the nervous system with a powerful mechanism for distinguishing meaningful signals from random background activity.
This filtering function is paramount for maintaining a high signal-to-noise ratio in complex circuits. Random, low-frequency inputs typically decay harmlessly because the inter-event intervals are too large to engage summation effectively. Only the temporally aligned, coherent signals indicative of a genuine event are granted access to the next stage of the neural pathway. This principle is fundamental to processes like acoustic perception, where the sustained, rapid input generated by a continuous tone is integrated via temporal summation to create the continuous perception of sound, rather than a series of discrete clicks. In motor systems, temporal summation is equally crucial. Sustained muscle contraction requires the motor neurons to fire continuously, which is achieved only when the central command provides a high-frequency burst of EPSPs that are temporally summed to maintain the postsynaptic membrane potential above the action potential threshold.
In higher-order cognitive functions, such as learning and memory, temporal summation contributes to synaptic plasticity. The induction of crucial memory mechanisms, such as Long-Term Potentiation (LTP), often requires the presynaptic neuron to fire at high frequencies (tetanic stimulation). This requirement for high-frequency input ensures that sufficient temporal summation occurs on the postsynaptic side, leading to a substantial and prolonged depolarization necessary to activate voltage-dependent receptors (like NMDA receptors) and trigger the biochemical cascade that strengthens the synapse. Thus, temporal summation acts as a gatekeeper, ensuring that only temporally robust and biologically relevant activity patterns are encoded and retained by the memory system.
Experimental Evidence and Measurement Techniques
The principles of temporal summation have been empirically verified through classic and modern electrophysiological methodologies. Early investigations relied heavily on intracellular recording techniques, primarily utilizing sharp microelectrodes inserted into the postsynaptic neuron to measure precise changes in membrane potential while stimulating the presynaptic axon at controlled frequencies. By delivering pairs of stimuli with varying inter-stimulus intervals (ISIs), researchers could graphically demonstrate the additive nature of PSPs. When the ISI was long, two separate, subthreshold PSPs were observed. As the ISI was systematically reduced to be shorter than the membrane time constant, the peak of the second PSP was observed to rise significantly higher than the first, confirming the summation effect. Furthermore, upon reaching a critical, very short ISI, the combined potential successfully generated an action potential, providing direct proof of the mechanism’s role in threshold determination.
Quantitative analysis of temporal summation often involves calculating the membrane time constant ($tau$) from the exponential decay curve of an isolated PSP, and then relating this value to the required stimulus frequency for efficient summation. Frequency-response curves are generated, plotting the amplitude of the summed potential against the stimulation frequency, clearly illustrating that the summation efficiency drops off dramatically when the frequency falls below $1/tau$. Modern patch-clamp techniques allow for exquisite control and measurement, enabling researchers to isolate specific ionic currents contributing to the PSP and its persistence. For instance, voltage-clamp protocols can isolate the total charge flow during a train of high-frequency stimuli, providing a quantitative measure of the accumulated charge that contributes to the summed depolarization.
The advent of optogenetics has provided a powerful tool for studying temporal summation in complex, living circuits. By genetically engineering specific presynaptic neurons to express light-sensitive proteins (opsins), researchers can precisely control the timing and frequency of action potentials using pulses of light. This level of temporal precision allows for the highly controlled induction of temporal summation trains, confirming its operation within intact neural pathways and allowing investigation into how summation efficiency might vary across different cell types and brain regions. These advanced techniques reinforce the understanding that temporal summation is a highly conserved and critical biophysical mechanism underlying neuronal computation across the central and peripheral nervous systems.
Clinical Relevance and Pathophysiology
The proper functioning of temporal summation is essential for neurological health, and disturbances to this mechanism can be implicated in various clinical disorders. Any condition that pathologically alters the membrane resistance or capacitance of neurons—thereby changing the time constant—can severely impair the neuron’s ability to integrate input correctly. For instance, channelopathies, which are disorders caused by mutations in ion channel genes, can lead to either excessively brief or excessively prolonged PSPs. If the PSP duration is too short, the capacity for temporal summation is reduced, potentially leading to hypoexcitability and failure to transmit necessary signals. Conversely, if the PSP is pathologically prolonged, the neuron may become hyperexcitable, leading to inappropriate firing patterns.
In conditions such as epilepsy, the hallmark is often neuronal hyperexcitability, characterized by synchronized, excessive firing. While multiple factors contribute, an increase in the effective time constant or a failure of inhibitory mechanisms can lead to uncontrolled temporal summation of EPSPs. Pharmacological treatments for epilepsy often target this balance, for example, by enhancing GABAergic inhibition (via drugs like benzodiazepines), which increases the magnitude and duration of IPSPs. These inhibitory potentials effectively counteract the excitatory drive, making it significantly harder for the temporal summation of EPSPs to reach the firing threshold, thus stabilizing the neuronal network and preventing seizures.
Furthermore, temporal summation is critical in managing sensory processing disorders. In conditions such as sensory overload, often observed in autism spectrum disorder or generalized anxiety, it is hypothesized that there may be an imbalance in the filtering capacity of the neural circuits. If the filtering mechanism—the ability to distinguish between signal and noise via efficient summation—is impaired, either through ineffective inhibition or overly excitable membrane properties, the brain may inappropriately amplify or over-integrate low-level sensory inputs, leading to the perception of overwhelming stimuli. Therefore, understanding and potentially therapeutically targeting the dynamic regulation of the membrane time constant offers a promising avenue for modulating neural excitability and restoring functional integration in a variety of neurological and psychiatric conditions.