Neuroanatomy: How Spinal Roots Shape Your Mental Reality
Introduction to the Spinal Root
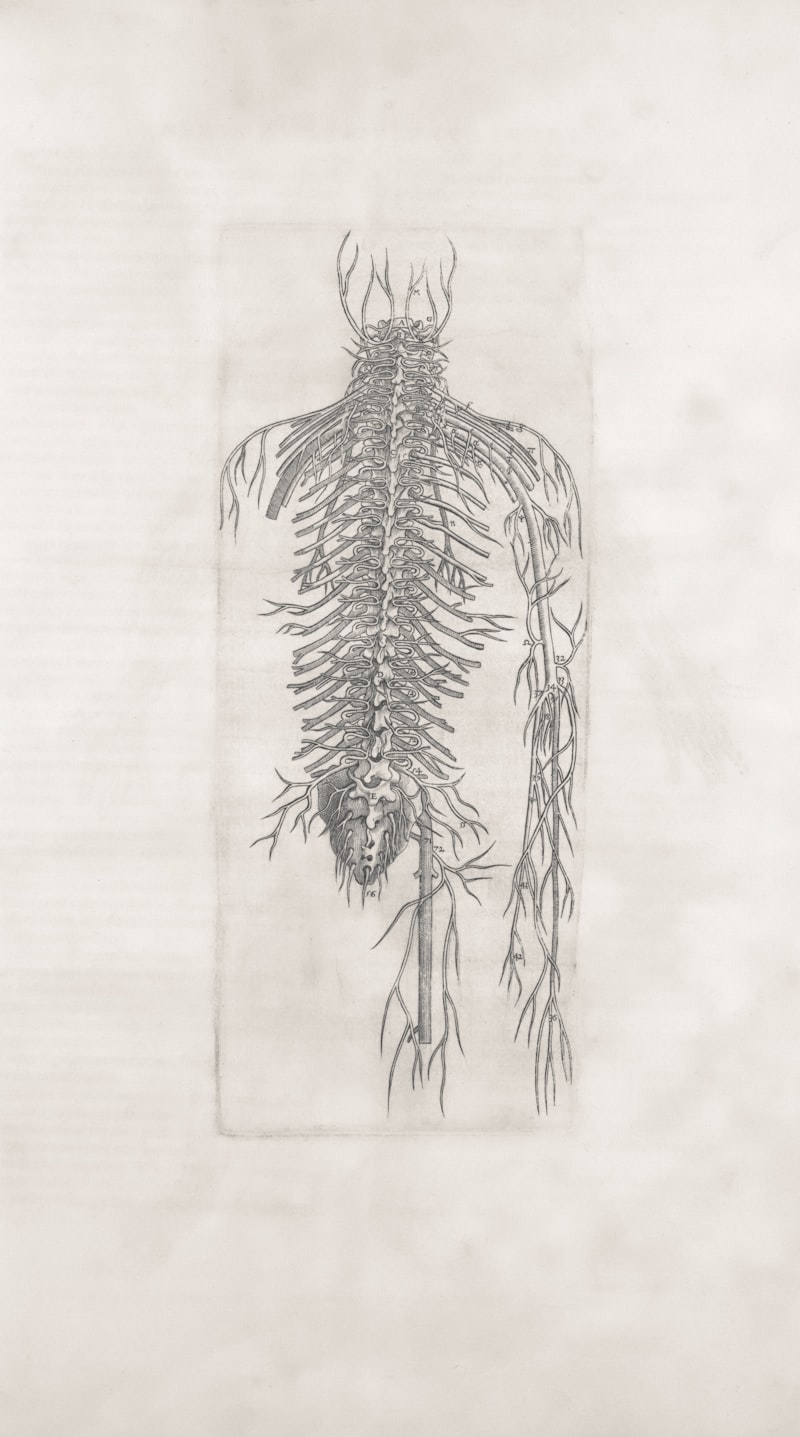
The spinal root represents a critical juncture in the organization of the human nervous system, serving as the essential anatomical and functional link between the central nervous system (CNS)—specifically the spinal cord—and the peripheral nervous system (PNS). This structure is not a singular entity but rather a complex bifurcation point where the signals traveling to and from the body are segregated based on their function, a principle foundational to neurophysiology. Each spinal nerve, which arises from the spinal cord, must first pass through this root structure, dividing emphatically into two distinct components: the dorsal root and the ventral root. This strict separation ensures that sensory information, which is derived from the external environment and internal bodily states, is channeled exclusively through one pathway, while motor commands, dictating muscle contraction and glandular secretion, are restricted entirely to the other. Understanding the spinal root is paramount for comprehending how bodily sensations are processed and how voluntary and involuntary movements are executed, highlighting its fundamental role in maintaining homeostasis and facilitating interaction with the environment.
The anatomical positioning of the spinal root is highly specific, located immediately adjacent to the spinal cord within the protective confines of the vertebral column. As the spinal cord extends caudally from the brainstem, it gives rise to thirty-one pairs of spinal nerves, each corresponding to a segment of the vertebral column (cervical, thoracic, lumbar, sacral, and coccygeal). Before the fusion point where the dorsal and ventral components merge to form the mixed spinal nerve, these root components exit the spinal cord via smaller filaments known as rootlets. These rootlets coalesce to form the larger dorsal and ventral roots, which then pass through the intervertebral foramina. This precise arrangement dictates that any structural compromise, such as a herniated disc or osteophyte formation narrowing the foramen, directly impinges upon these delicate root structures, leading to predictable neurological deficits known collectively as radiculopathy. The integrity of the spinal roots is thus inseparable from the overall health and function of the musculoskeletal and nervous systems.
The functional dichotomy established by the spinal root is one of the most elegantly simple yet powerful organizational principles of neural architecture. The dorsal root is dedicated entirely to afferent signaling—that is, transmitting sensory input toward the CNS. This input encompasses a wide array of modalities, including pain, temperature, touch, proprioception (body position), and vibration. Conversely, the ventral root is the conduit for efferent signaling, carrying motor commands away from the CNS to effector organs, primarily skeletal muscles for voluntary movement, but also including autonomic fibers directed toward smooth muscles and glands. This clear segregation of function (the Bell-Magendie Law) is essential for rapid and accurate signal processing, preventing the confusing overlap of incoming sensory data and outgoing motor commands at the critical junction point near the spinal cord grey matter.
Anatomical Foundation and Location
The architecture supporting the spinal roots is robust, yet highly vulnerable due to its close proximity to the bony structures of the vertebral column. Each pair of spinal roots emerges laterally from the spinal cord, specifically within the dura mater, and traverses the subarachnoid and epidural spaces before uniting just distal to the dorsal root ganglion (DRG) to form the main spinal nerve trunk. The roots themselves are composed of hundreds of individual myelinated and unmyelinated axons bundled together by connective tissue sheaths. The precise point of fusion between the dorsal and ventral roots occurs just medial to the intervertebral foramen. Once the fusion is complete, the resulting spinal nerve immediately branches again into the dorsal ramus (supplying the deep back muscles and skin) and the ventral ramus (supplying the limbs and the anterior trunk), demonstrating the rapidity with which peripheral distribution begins after CNS exit.
The arrangement of the rootlets is highly organized, reflecting the segmentation of the spinal cord itself. The ventral rootlets emerge from the ventrolateral sulcus of the spinal cord, originating from motor neuron cell bodies located within the ventral horn of the grey matter. In contrast, the dorsal rootlets enter the spinal cord at the posterolateral sulcus. This spatial separation within the grey matter—motor nuclei anteriorly and sensory relay nuclei posteriorly—is maintained throughout the root structure. Furthermore, the length and orientation of the spinal roots vary significantly depending on their level of origin. The cervical roots are relatively short and horizontal, whereas the lumbar and sacral roots descend almost vertically within the vertebral canal, forming the bundle known as the cauda equina (horse’s tail) before exiting their respective foramina. This caudal elongation makes these lower roots particularly susceptible to injury within the confined space of the lower spine.
A key anatomical feature intrinsically associated with the dorsal root is the dorsal root ganglion (DRG). This ovoid enlargement is situated along the dorsal root just proximal to its merger with the ventral root. The DRG houses the cell bodies of all primary sensory neurons. These neurons are typically pseudounipolar, meaning they have a single process that splits shortly after leaving the cell body, one branch extending peripherally to receive sensory stimuli and the other extending centrally to enter the spinal cord. The presence of the DRG confirms the exclusively sensory nature of the dorsal root, as the cell bodies for motor neurons are located safely within the ventral horn of the spinal cord grey matter. This architectural separation underscores the protective mechanism that shields the vital motor centers from peripheral damage that might primarily affect the sensory ganglia.
The Dorsal Root: Afferent Pathway
The primary function of the dorsal root is the transmission of afferent signals, initiating the communication pathway from the external environment and the internal viscera to the CNS for processing. These sensory fibers originate from receptors distributed throughout the skin, muscles, tendons, joints, and organs. The information carried is diverse, ranging from highly specific fine touch and proprioceptive data, which travels quickly along heavily myelinated A-beta fibers, to slower, more diffuse signals related to crude touch, temperature, and pain, often carried by thinly myelinated A-delta fibers and unmyelinated C fibers. The sheer volume and complexity of the sensory data funneled through the dorsal roots necessitate highly organized tracts within the spinal cord for subsequent ascent to higher brain centers, such as the thalamus and somatosensory cortex.
The organization of sensory input via the dorsal roots follows a predictable pattern mapped onto the body surface known as dermatomes. Each of the thirty-one pairs of dorsal roots is responsible for collecting sensory information from a specific, relatively defined area of the skin. This segmental organization is immensely valuable in clinical neurology; for instance, pain or numbness localized to a specific dermatome (e.g., pain wrapping around the chest at T4) strongly suggests a lesion or compression affecting that particular spinal root rather than a peripheral nerve injury, which typically follows the distribution of the mixed nerve distal to the root origin. The precise mapping allows clinicians to localize the level of spinal cord or root injury, demonstrating the direct relationship between the anatomical structure of the root and the physiological perception of sensation.
Once the sensory fibers enter the spinal cord via the dorsal root, they have several critical destinations and functional roles. Some fibers ascend immediately within the dorsal columns (the Fasciculus Gracilis and Cuneatus) to relay precise touch and proprioception directly to the brainstem. Other fibers synapse almost immediately within the dorsal horn of the grey matter, participating in local reflexes. For example, nociceptive (pain) fibers synapse with interneurons that can quickly activate motor neurons via the ventral root to initiate withdrawal reflexes. Furthermore, crucial relay neurons in the dorsal horn modulate incoming pain signals before they ascend, a process vital for pain management and understanding conditions of chronic pain sensitization. Thus, the dorsal root is not merely a passive cable; it initiates complex processing immediately upon entering the CNS.
The Ventral Root: Efferent Pathway
The ventral root is the exclusive carrier of efferent signals, transporting motor commands originating within the spinal cord out to the periphery. These signals are the execution arm of the nervous system, responsible for all voluntary movement, muscle tone maintenance, and a significant portion of autonomic function. The axons comprising the ventral root originate from the large, multipolar cell bodies of motor neurons situated within the anterior (ventral) horn of the spinal cord grey matter. These neurons are categorized primarily as alpha motor neurons, which directly innervate and cause the contraction of skeletal muscle fibers, and gamma motor neurons, which innervate the specialized muscle fibers within muscle spindles, thereby regulating muscle sensitivity.
The motor output channeled through the ventral root is highly organized somatotopically. Motor neurons innervating proximal muscles (e.g., shoulder and hip) are generally located more medially within the ventral horn, while those innervating distal muscles (e.g., hand and foot) are located more laterally. This precise mapping ensures that commands for complex, coordinated movements are delivered synchronously and accurately. Damage to the ventral root directly results in flaccid paralysis of the muscles it supplies, as the necessary excitatory input from the CNS is completely severed, leading to muscle atrophy over time. This contrasts sharply with upper motor neuron lesions, which often cause spasticity because the spinal reflexes themselves remain intact but are released from descending inhibition.
In addition to somatic motor fibers supplying skeletal muscles, the ventral roots at specific spinal levels (T1-L2 and S2-S4) also carry preganglionic autonomic fibers. The thoracic and upper lumbar ventral roots carry preganglionic sympathetic fibers, which exit the cord and travel to the sympathetic chain ganglia, regulating functions like heart rate, vasoconstriction, and glandular secretion. The sacral ventral roots carry preganglionic parasympathetic fibers, which control pelvic visceral functions such as bladder and bowel emptying. Therefore, while often simplified as purely the motor pathway, the ventral root is functionally heterogeneous, encompassing both the immediate control of movement and the critical regulation of involuntary visceral functions necessary for survival.
Functional Significance in Reflex Arcs
The structure of the spinal root is fundamentally optimized for the execution of spinal reflex arcs, the rapid, involuntary responses that are vital for protection and posture maintenance. A basic reflex arc requires an uninterrupted circuit: sensory input travels via the dorsal root, synapses within the spinal cord grey matter (either directly onto a motor neuron or via an interneuron), and the resulting motor command exits via the ventral root. This entire process bypasses conscious brain processing, allowing reaction times measured in milliseconds. The physical separation of the dorsal (in) and ventral (out) roots ensures that the flow of information for the reflex is unidirectional and efficient.
The simplest example is the monosynaptic stretch reflex, such as the knee-jerk reflex. When the patellar tendon is tapped, sensory afferents in the muscle spindle activate. These signals travel through the dorsal root and enter the spinal cord, where they synapse directly onto the alpha motor neurons in the ventral horn. The motor command then exits immediately via the ventral root, causing the quadriceps muscle to contract. The integrity of both the dorsal and ventral roots at the specific spinal segment (L2-L4 for the patellar reflex) is absolutely necessary for a normal response. If either root is damaged, the reflex will be diminished or absent, a key clinical indicator of root pathology.
Furthermore, the spinal roots facilitate more complex, polysynaptic reflexes, which involve one or more interneurons within the spinal cord. The withdrawal reflex, for example, involves sensory input (pain) entering the dorsal root, synapsing with interneurons that simultaneously excite motor neurons for the flexor muscles (to pull the limb away) and inhibit motor neurons for the extensor muscles (to prevent opposing movement). This coordination, known as reciprocal innervation, is entirely dependent on the precise segregation and rapid communication provided by the dorsal root supplying the sensory information and the ventral root executing the complex, coordinated motor response across multiple muscle groups.
Clinical Relevance and Pathologies
Pathologies affecting the spinal roots, collectively termed radiculopathies, constitute a significant portion of clinical neurological practice. Because the roots are enclosed within the bony confines of the vertebral canal and exit through the narrow intervertebral foramina, they are highly susceptible to mechanical compression and inflammation. The most common cause of radiculopathy is the herniation or protrusion of an intervertebral disc, which impinges directly upon the root structure as it attempts to exit the foramen. Since the sensory and motor fibers are tightly bundled together in the pre-fusion root, compression often results in a mixed presentation of symptoms.
Symptoms of radiculopathy typically include sensory deficits corresponding to the affected dermatome, manifesting as pain (radicular pain, often shooting), numbness, tingling (paresthesia), or heightened sensitivity (dysesthesia). Motor involvement, due to compression of the ventral root fibers, presents as weakness, muscle wasting, and diminished or absent deep tendon reflexes corresponding to the affected segment. For example, L5 radiculopathy commonly causes weakness in ankle dorsiflexion (foot drop) and sensory changes along the lateral leg and top of the foot. Accurate diagnosis requires understanding the precise segmental distribution of both sensory and motor function dictated by the specific root involved.
Beyond mechanical compression, spinal roots can be affected by infectious and inflammatory processes. Viral infections, particularly the Varicella Zoster Virus (which causes shingles), often lie dormant within the sensory neuron cell bodies of the dorsal root ganglia (DRG). Reactivation of the virus leads to inflammation and destruction of the DRG and the corresponding dorsal root fibers, causing severe, characteristic radicular pain and a vesicular rash distributed precisely along the affected dermatome. Furthermore, autoimmune conditions such as Guillain-Barré Syndrome can involve inflammation of the roots themselves (polyradiculopathy), leading to rapidly progressive weakness and paralysis, demonstrating that the spinal roots are vulnerable not only to external compression but also to systemic disease processes.
Development and Embryology
The formation of the spinal roots is a complex process intertwined with the early development of the neural tube and the adjacent mesoderm. The spinal cord itself differentiates from the neural tube, establishing the alar plate (future dorsal horn) and the basal plate (future ventral horn). This initial segregation determines the functional destiny of the fibers that will eventually form the roots. The motor components of the ventral root develop early, as motor neuroblasts migrate and differentiate within the basal plate, extending their axons ventrally to exit the spinal cord and form the ventral rootlets, which seek out their target muscles in the developing limb buds and trunk musculature.
The sensory components of the dorsal root originate from an entirely separate structure: the neural crest cells. These pluripotent cells migrate laterally from the closing neural tube and aggregate to form the primordial dorsal root ganglia (DRG). The neurons within the DRG differentiate into the pseudounipolar sensory neurons. Crucially, these neurons must extend two processes simultaneously: a central process that grows inward to enter the dorsal horn of the developing spinal cord, thereby forming the dorsal root, and a peripheral process that grows outward to innervate peripheral receptor fields. This dual origin—motor fibers arising from the CNS basal plate and sensory fibers arising from the PNS neural crest—reinforces the organizational separation maintained throughout adult life.
Errors during the embryonic development of the spinal roots can lead to congenital anomalies. Conditions such as spinal dysraphism (e.g., spina bifida) often involve malformation or tethering of the spinal cord and its roots due to incomplete closure of the neural arch. In such cases, the abnormal traction or compression placed on the developing dorsal and ventral roots can result in severe motor and sensory deficits, particularly affecting the lower limbs and bladder function. The proper chronological and spatial alignment of motor axon outgrowth and sensory neuron migration is therefore essential for establishing the functional integrity of the spinal root system.
Relationship to the Peripheral Nervous System (PNS)
The spinal root occupies a unique transitional zone, representing the immediate peripheral extension of the CNS before its structures fully merge into the PNS. The fundamental distinction between the CNS and PNS often relies on the location of neuronal cell bodies and axons: tracts within the CNS are typically myelinated by oligodendrocytes, while nerves in the PNS are myelinated by Schwann cells. The spinal root is the point where this transition occurs. The axons of the ventral root are still technically within the CNS environment as they exit the cord, but once they pierce the pia mater and are surrounded by Schwann cells, they become part of the PNS.
The dorsal root ganglion (DRG), despite being closely associated with the spinal cord, is definitively a component of the PNS because the cell bodies of the sensory neurons reside outside the CNS. This placement is strategically important: if the peripheral process of a sensory neuron is damaged, the cell body in the DRG often has a greater capacity for regeneration than if it were located within the spinal cord itself. However, the central processes of these DRG neurons, which form the dorsal root, must successfully regenerate into the hostile CNS environment if damaged near the spinal cord, highlighting the complex regenerative challenges posed by injury at the root level.
In summary, the spinal root system serves as the gateway through which all communication between the brain, spinal cord, and the rest of the body must pass. The formation of the spinal nerve, resulting from the perfect anatomical fusion of the functionally specialized dorsal and ventral roots, marks the formal beginning of the peripheral nervous system’s vast network. The integrity of this junction is paramount, as demonstrated by the profound motor and sensory deficits that arise when the roots are compromised, confirming their status as essential conduits for neurological function.